Nonlinear dynamics and stochastic methods: from neuroscience to other biological applications
Conference venue:
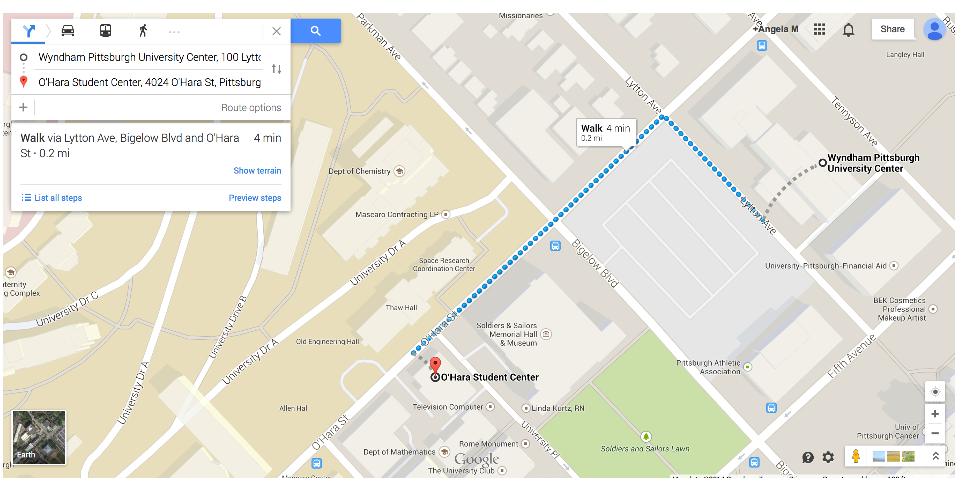
O'Hara Student Center, University of Pittsburgh
(see map)
3900 O'Hara Street, Pittsburgh, PA 15260
Note: Wyndham Hotel is very close to O'Hara Student Center (see map)
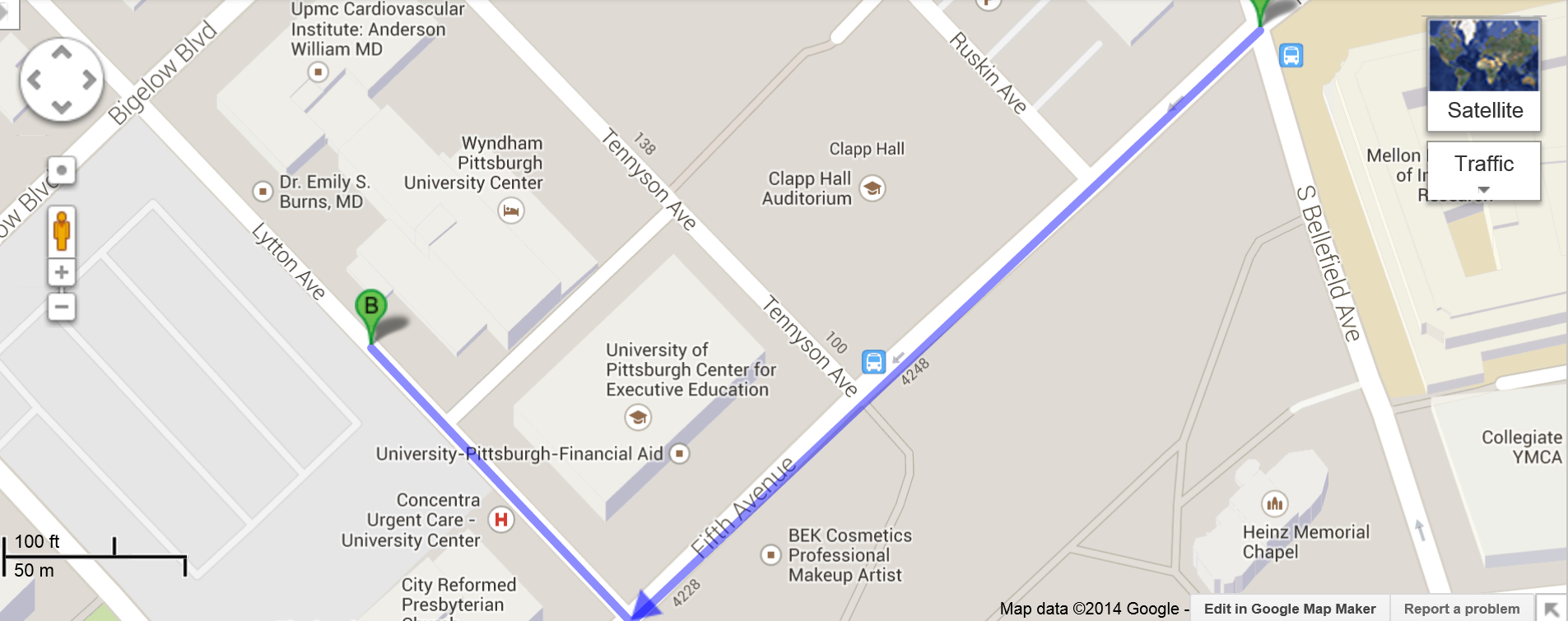
Easy transportation from the airport to the hotel by bus: take 28X Airport Flyer (fare: $3.75 - exact change is required); get off at stop S. Bellefield Ave at Fifth Avenue (in Oakland); then walk 4 min to Wyndham Hotel
March 10-12, 2014
University of Pittsburgh - Pittsburgh, PA
In conjunction with the conference, we plan to honor Professor Bard Ermentrout who has been at the forefront of the conference's topics throughout his career (see conference flier).
Organizers
Rodica Curtu, University of Iowa
Angela Reynolds, Virginia Commonwealth University
Local Committee
Brent Doiron, University of Pittsburgh
Jonathan Rubin, University of Pittsburgh
This three-day conference will bring together a mix of senior and junior scientists
to report on theoretical methods that proved successful in mathematical neuroscience,
and to encourage their dissemination and application to modeling in computational medicine
and other biological fields. The invited speakers will present on mathematical topics such
as dynamical systems, multi-scale modeling, phase resetting curves, pattern formation and
statistical methods.
The mathematical tools will be demonstrated in the context of the following main topics:
1. Rhythms in biological systems;
2. The geometry of systems with multiple time scales;
3. Pattern formation in biological systems;
4. Stochastic models: statistical methods and mean field approximations.
Participants
List of confirmed participants
Invited Speakers
| Paul Bressloff | University of Utah | ||
| Carson Chow | National Institutes of Health | ||
| Jack Cowan | University of Chicago | ||
| Sharon Crook | Arizona State University | ||
| Lance Davidson | University of Pittsburgh (Bioengineering) | ||
| Jonathan Drover | Cornell Medical College | ||
| Leah Edelstein-Keshet | University of British Columbia | ||
| Roberto Fernandez Galan | Case Western Reserve University | ||
| Pranay Goel | Indian Institute of Science, Education and Research, Pune - India | ||
| Boris Gutkin | Ecole Normale Superieure, Paris - France | ||
| Zachary Kilpatrick | University of Houston | ||
| Nancy Kopell | Boston University | ||
| Cheng Ly | Virginia Commonwealth University | ||
| Remus Osan | Georgia State University | ||
| John Rinzel | New York University | ||
| Jonathan Rubin | University of Pittsburgh (Mathematics) | ||
| Daniel Simons | University of Pittsburgh (Neurobiology) | ||
| David Terman | Ohio State University |
Program and Abstracts
Conference schedule
Abstracts of confirmed poster presentations
Maximum dimensions for posters: 48 inches (height) x 46 inches (width)
Abstracts of the invited talks:
Paul Bressloff (University of Utah) Breakdown of fast-slow analysis in an excitable neuron with channel noise Abstract  | |
Carson C. Chow (National Institutes of Health) How many neurons code a percept? Abstract | |
Jack Cowan (University of Chicago) Geometric Visual Hallucinations: what they tell us about the architecture of the brain Abstract | |
Sharon Crook (Arizona State University) A continuum model approach for exploring the role of neuronal structure Abstract | |
Lance Davidson (University of Pittsburgh) Engines of cell shape change: actomyosin dynamics within the cell cortex Abstract | |
Jonathan Drover (Cornell Medical College) A mean-field model suggests a novel EEG analysis technique to index thalamocortical dynamics Abstract | |
Leah Edelstein-Keshet (University of British Columbia) From actin assembly to cell motility, with a little help from my friend Abstract | |
Roberto Fernandez Galan (Case Western Reserve University) Stochastic Neural Dynamics and Information Processing in the Autistic Brain Abstract | |
Pranay Goel (Indian Institute of Science, Education and Research, Pune - India) Using the Dual Oscillator Model (DOM) to study bursting in pancreatic islets Abstract | |
Boris Gutkin (Ecole Normale Superieure, Paris - France) Working with Gamma, Theta, Alpha Oscillations (and Noise Correlations) to Make Working Memory Work Abstract | |
Zachary Kilpatrick (University of Houston) Getting the most out of bumps Abstract | |
Nancy Kopell (Boston University) Brain Rhythms: Multiple Roles of Inhibition Abstract | |
Cheng Ly (Virginia Commonwealth University) Networks of heterogeneous neural oscillators Abstract | |
Remus Osan (Georgia State University) Targeting performances for stochastic models of neural growth with uniform branching and pruning Abstract | |
John Rinzel (New York University) Biased competition for context-dependent perceptual choice Abstract | |
Jonathan Rubin (University of Pittsburgh) ...and Out Come the Boundary Conditions Abstract | |
Daniel J. Simons (University of Pittsburgh) Receptive field transformations in feedforward thalamocortical circuits Abstract | |
David Terman (Ohio State University) What do small toy models tell us about large complicated networks? Abstract |

{kind=link}
{kind=link}
Announcements
The general goal of the conference is to present and demonstrate both the successes and challenges of mathematical modeling in neuroscience, and to encourage the dissemination and application of such techniques to modeling in other biological fields. Therefore, the organizing committee will strive to balance the number of neuroscience-related posters with posters presenting mathematical and computational results for other biological problems, such as disease dynamics, epidemiology and physiology (please see abstract submission). Special attention will be paid when selection the
poster presentations, priority will be given to young researchers, including graduate students, women and minorities.
Researchers interested in attending the conference should submit a registration application. The attendees will be selected among these based on research interest overlapping with the conference theme, and to ensure diversity and breadth of participation by individuals and institutions.
To ensure more general mathematical and computational
biology audience, the invited speakers will be asked to assume no prior knowledge of any particular biological area, when preparing their talk.
Important Deadlines
!!! NEW: Deadline for registration application is January 14th, 2014 at 11:59 PM EST !!!
September 10, 2013: Registration application and travel award application open.
Abstract submission open.
December 1, 2013: Travel award application close.
Abstract submission close.
!!! Extended deadline for abstract submission and travel award application: December 22, 2013 !!!
!!! Given the wide interest shown in the conference, we reopen the abstract submission and travel award application with FINAL DEADLINE: January 2, 2014. NOTE: The website may still accept submissions after the above date, but those received after 11:59 PM EST on 1/2/2014 will not be considered !!!
January 17, 2014: Authors notified about outcome of review
(Poster presentation: accept/reject).
(Registration: accept/reject).
Travel award notifications.
Registration Information / Travel Award Application
Please complete the registration application online by providing your name, home institution, position, research interest and email address.
Registration Application
Deadline for registration application (January 14th, 2014 at 11:59 PM EST) has passed!!!
Travel awards for students, postdocs and other early career researchers presenting a poster may become available. If you are interested in being considered for a travel award, you will have an opportunity to apply via the registration application. In order to complete the travel award application, you will need to email a copy of your CV. Please have it available.
Travel Award Application Deadline for travel award application has passed!!!
Submissions
The conference will start Monday 3/10/2014 in the morning and will end Wednesday 3/12/2014 at noon. There will be four 40-minute long invited talks each morning and three invited talks each afternoon.
There will be no parallel sessions, and all lectures will be held in the same conference hall.
A two-hour poster session is scheduled for Monday evening. Selected poster presenters will be notified by January 17, 2014 .
Abstract Submission Deadline for abstract submission has passed!!!
Hotel Information
Wyndham Pittsburgh University Center
100 Lytton Avenue Pittsburgh, PA 15213
A group rate ("Nonlinear Dynamic") is available for attendees to this conference, for the interval Sunday March 9th (check in) through Thursday March 13th (check out). The rate is $135/night + taxes. Parking is an additional charge of $14/day. The hotel is in walking distance to the conference site.
Hotel reservation deadline for this group rate: Sunday February 16th, 2014, via the following website:
Wyndham Pittsburgh University Center
Group code: 03096841ND (enter this code in the special rate code marked group code)